In Depth
Out of all the hadrosaurs (also known as the duck-billed dinosaurs) Parasaurolophus is one of the most widely recognised thanks to its very distinctive skull crest.
The name Parasaurolophus is a bit of a mouthful, but this is based around the early interpretation that Parasaurolophus was similar to another genus names Saurolophus (which means ‘lizard crest) because Saurolophus also has a skull crest, though not as large or as ornate as Parasaurolophus.
However under modern systematics, Parasaurolophus is classed as a lambeosaurine hadrosaurid because of the hollow crest, whereas Saurolophus is the type genus of the Saurolophinae (previously Hadrosaurinae) a sister group of hadrosaurids noted for having solid to no crests at all.
This means that Parasaurolophus and Saurolophus are in fact distantly related, but both are still called hadrosaurids because they both belong within the Hadrosauridae.
Instead Parasaurolophus is more closely related to other lambeosaurines such as Hypacrosaurus, Corythosaurus and of course the type genus of the group, Lambeosaurus. O
ut of all lambeosaurines however, it is the Asian genus Charonosaurus that is thought to be one of the closest relatives of Parasaurolophus due to the strikingly similar head crest.
The three main Parasaurolophus species
P. walkeri
This is the type species of Parasaurolophus, which means that the fossils that belong to this species are the ‘benchmark’ that all future fossil discoveries are compared to before they are formally added to the genus.
Named by William Parks in 1922, P. walkeri has been established upon a holotype (specific fossils used for identifying future discoveries) of a skull and partial skeleton recovered from what is now the Dinosaur Park Formation of Alberta, Canada.
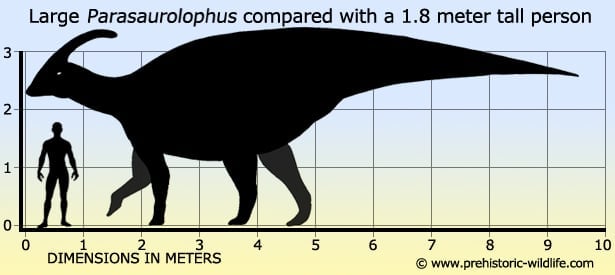
Although not a hundred per cent complete, this specimien has come from an individual that is estimated to have been around nine and a half meters long in life.
The skull of P. walkeri features the distinctive long arcing crest that rises up from the skull, but a more simplified network of air passages within. Apart from Canada, there has also been past speculation that P. walkeri remains may also be in Montana, USA, although so far not much has come of this.
P. tubicen
This is the second species of Parasaurolophus, first named in 1931 by Carl Wiman. The species name means ‘trumpeter’.
Externally the crest of P. tubicen is very much like that of P. walkeri, but the nasal passages inside are more complex in form.
P. tubicen seems to have grown slightly larger than P. walkeri, and is so far known from the remains of at least three individuals.
However it is still hard to establish the size of P. tubicen in relation to P. walkeri since the two species together are only represented by fossils for a few individuals, but hopefully future discoveries may allow for a more complete picture.
So far P. tubicen is best known from Kirtland Formation and possibly the Fruitland Formation of New Mexico, USA.
P. cyrtocristatus
Named in 1961 by John Ostrom, this is an interesting species of Parasaurolophus and one destined for a lot of future study.
The crest of this species is much shorter than the crests of P. walkeri and P. tubicen, and also has a stronger curvature to the point where the end actually reaches around the back of the skull. P. cyrtocristatus also seems to be the smallest known species.
Although still widely accepted as being a valid species, study of other hadrosaurids, particularly lambeosaurines has led to the idea that P. cyrtocristatus might actually be juvenile P. tubicen, or alternatively females of P. tubicen.
This would actually fit into other patterns set by other genera where specimens interpreted as female seem to have slightly different and reduced crests to those thought to be male.
The main counter to this idea however comes from the simple observation that P. cyrtocristatus appears about one million years before P. tubicen.
This does not negate the possibility that the two species are the same, but ideally remains of both species need to be found in the same time zones, preferably together. P. cyrtocristatus is currently known from the Kaiparowits Formation of Utah, USA.
What did Parasaurolophus use its head crest for?
There have been many theories about what Parasaurolophus used its skull crest for which has led to many erroneous collections of facts about this dinosaur, but there are three theories in particular that have strong merit.
The first theory is that of visual display, an idea that explains the difference in crest shape as being a signature feature between different species.
A possible scenario would be an ecosystem where more than one species or genus of lambeosaurine hadrosaur was present, but it was difficult to tell the difference between them because of the similar size and body forms between them.
One look at the head crest however would allow for instant recognition between species so that they could then focus upon attracting and pairing up with their own species.
The second theory that is actually linked to the first is that of auditory communication.
The crest of Parasaurolophus houses tubes that run up from the skull to the tip where they curve round and run back down to the skull.
This has given rise to the resonating chamber theory where calls from the throat of the dinosaur pass through the passages where they are amplified by the crest so that they are louder and have a broader frequency.
Because lambeosaurine crests are usually quite different between species and genera, the different shapes would produce different levels of amplification, in short creating different sounding calls distinct and unique to a species.
Additionally going back to the idea that male and female Parasaurolophus may have had slightly different shaped crests, the males might have been more suited for producing louder and longer calls.
Just like how male deer (bucks) roar to signal their strength, the louder and longer calls of Parasaurolophus (and other lambeosaurines) could have belonged to the healthiest and possibly most mature males, both traits desired by females.
Further support for the auditory theory comes from study of lambeosaurine hearing systems (particularly those of Corythosaurus) which suggest that hearing was one of the most important and developed senses in lambeosaurine hadrosaurids.
The idea of auditory calling has received a lot of attention for lambeosaurines, but Parasaurolophus in particular has been the most popular study, including computer reconstructions of the crest with what the sound might have been like when air was passed through.
The third contender is that of thermoregulation, particularly with cooling for the skull and brain. First proposed in 1978 by P.E.
Wheeler, the idea is based around the crest providing a larger surface area for heat to exchange through.
This is a simple principal where say the only factor involved for losing heat that is changed is the surface area, so for the sake of argument, a ten square centimetre area should lose heat ten times faster than a one centimetre area because of the increased surface area.
Other clues for thermoregulation come from the skeleton where the high neural spines of the vertebrae significantly increase the height and surface area of the body for a much smaller proportionate increase in mass.
Unfortunately the problem with the thermoregulation theory is that it does not explain the difference in crest forms for other lambeosaurine genera.
If the reason for the crest was more biological, then it would make more sense if the crests all developed along similar lines with little variation.
It is quite possible however that more than one of the above theories is correct, after all there is no reason to assume that a physical feature can only have one purpose.
Additionally the hollow crest may have originally come about as a weight saving feature to reduce skull weight before adapting to include another purpose.
Older theories do of course resurrect themselves from time to time, so let’s take a brief look at these.
Head/Neck support
Actually proposed by William Parks, the palaeontologist who first described Parasaurolophus.
The idea is that the crest served as a muscle attachment point for stabilising the head and neck.
However there are no signs of muscle attachment points on the neck, and even if the crest was for this function, it would probably hinder this dinosaurs ability to move its neck.
Also does not explain the variance in other lambeosaurines since they seem to have managed just fine without any extra support.
Air trap
Proposed by Charles M. Sternberg to keep water out of the lungs, but no feasibly practical way has been produced to demonstrate how which is why it’s considered unlikely.
In a variation Ned Colbert suggested that it could be an air reservoir used for when the head is underwater.
This is considered unlikely due to the very small storage capacity of the crest, and in such a situation Parasaurolophus would have probably been better off just taking a deep breath.
Snorkel
By Alfred Sherwood Romer, the idea is that Parasaurolophus breathed through the crest so that it did not have to lift its head above the surface of the water.
Obvious flaw here is that the crest is not a tube, the end is solid bone. This makes the snorkel idea impossible.
Weapon
By Othenio Abel, but the crest does not have an especially robust construction, and weapons tend to face outwards rather than inwards towards the body.
The only feasible way this could work is if a Parasaurolophus dipped its head down and then flicked it up so that the end of the crest struck in an uppercut motion.
Attachment of a Proboscis
By Martin Wilfarth, a proboscis is basically similar to the trunk of an elephant.
Why a proboscis would attach to the back of the head is still unknown however since all other vertebrates known to have these always have them on the front.
Branch guard
By Andrew Milner, but this depends upon what Parasaurolophus ate.
If Parasaurolophus was hyper specialised in its diet and only ate low growing vegetation that grew under taller and denser vegetation then this might have a benefit.
Otherwise it seems strange that a herbivorous dinosaur should have an adaptation for holding away plants it was eating.
Storage of salt glands
Proposed by Halszka Osmolska, and probably the most unique and interesting one here.
Salt glands exist to excrete excessive amounts of salts that have been absorbed from the surrounding environment, however they are usually only found upon marine creatures, that is animals that either live in or next to the sea.
There is no definitive evidence that Parasaurolophus lived in marine environments, though their known fossil locations of Alberta and New Mexico indicate that they did live fairly close to the Western Interior Seaway, a shallow Cretaceous sea that submerged most of central North America from what is now the Arctic Ocean all the way down to the Gulf Mexico.
The idea is considered tentative at best as it does not explain why an unusual crest was needed for this, or indeed the differences between separate species.
Nasal capacity for a greater sense of smell
By John Ostrom and quite a logical proposal given the hollow construction of the crest and its close proximity to the nasal cavity.
However unless Parasaurolophus specialised in eating hidden plant parts like those below ground, there is no evidence to suggest that Parasaurolophus did this (i.e. specialised digging hands).
Sense of smell might have enabled Parasaurolophus to better detect predators, but predators usually try to approach from downwind of their prey, behaviour that renders the sense of smell next to useless.
Also does not explain why saurolophine hadrosaurids that lacked such elaborate had ornamentation seem to have replaced lambeosaurines in North America by the end of the Cretaceous as the dominant group of hadrosaurids.
Parasaurolophus as a dinosaur of the late Cretaceous
Once you get past the head crest, Parasaurolophus was pretty much just like every other lambeosaurine hadrosaurid.
Most of the time the body weight would have been supported on all four limbs, though Parasaurolophus probably could still get about quite competently on just the rear legs.
This ability to switch between postures meant that Parasaurolophus like other hadrosaurids had the flexibility to adapt to feeding at different heights from low down near the ground to reaching up into the lower tree canopy.
Additionally as a lambeosaurine, Parasaurolophus had a slightly narrower mouth than saurolophine hadrosaurs, suggesting that it could have been a more selective browser.
There is fossil evidence to strongly suggest that hadrosaurid dinosaurs like Parasaurolophus were prey to the top predators of the Campanian, principally the tyrannosaurs.
In the north around Canada and northern portions of the USA the main genera that could have posed a serious threat to adult Parasaurolophus would have been Albertosaurus, Gorgosaurus and Daspletosaurus.
Albertosaurus and Gorgosaurus in particular have a more gracile (lightweight) build which means that they would have likely been faster than others such as Daspletosaurus.
This would have given these two genera a serious edge in hunting hadrosaurids which could have been some of the fastest ‘large’ herbivores in the ecosystem, especially when you compare them to the horned ceratopsian dinosaurs like Chasmosaurus and armoured ankylosaurs like Euoplocephalus.
Further south and the slightly shorter snouted tyrannosaurs such as Bistahieversor and Teratophoneus were the main threats to the more southern species of Parasaurolophus.
Smaller predators however should not be discounted as threats, especially against smaller juveniles of Parasaurolophus.
One strong contender is that of Troodon, a dinosaur that although small when compared to a tyrannosaur, could have combined keen eyesight, slicing teeth and perhaps most importantly of all, intelligence to bring down smaller individuals.
Further Reading
- - Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur, William A. Parks - 1922.
- - On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks, Charles W. Gilmore - 1924.
- - Parasaurolophus tubicen n. sp. aus der Kreide in New Mexico [Parasaurolophus tubicen n. sp. from the Cretaceous in New Mexico], Carl Wiman - 1931.
- - A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico, John H. Ostrom - 1961.
- - The cranial crests of hadrosaurian dinosaurs, John H. Ostrom - 1962.
- - The evolution of cranial display structures in hadrosaurian dinosaurs, James A. Hopson - 1975.
- - Parasaurolophus (Reptilia: Hadrosauridae) from Utah, David B. Weishampelp, James A. Jenson - 1979.
- - Aspects of hadrosaurian cranial anatomy, Teresa Maryanska & Halszka Osmolska - 1979.
- - The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia), David B. Weishampel - 1981.
- - Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia:Ornithischia), David B. Weishampel - 1981.
- - A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications, Robert M. Sullivan & Thomas E. Williamson - 1996.
- - A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen, Carl F. Diegert & Thomas E. Williamson - 1998.
- - A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, Robert M. Sullivan & Thomas E. Williamson - 1999.
- - A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus, David C. Evans, Robert R. Reisz & Kevin Dupuis - 2007.
- - An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta, D. C. Evans, R. Bavington & N. E. Campione - 2009.