In Depth
Edmontosaurus is without a doubt one of if not the most intensively studied hadrosaurid dinosaur. Not only are there many Edmontosaurus specimens known, but those that have been recovered include some of the best preserved hadrosaurid material so far found.Classification history
Edmontosaurus has one of the longest and most complicated taxonomic histories of any dinosaur, and it is a subject that would need an entire article in its own right to explain properly. The first Edmontosaurus fossils were being found at least as far back as the late nineteenth century during a period in American paleontological history known as the Bone Wars. During this period two rival palaeontologists, Edward Drinker Cope and Othniel Charles Marsh, were desperately trying to outdo one another by naming as many new prehistoric animals (including but only dinosaurs) that they could. This led to a great many genera and species being created quickly without adequate study, and the task of identifying them properly fell to later palaeontologists.
The first Edmontosaurus fossils to be named were by Edward Drinker Cope in 1871, but Cope named the remains as belonging to a species of Trachodon (today regarded as a dubious genus of hadrosaur), T. atavus. Later in 1892, Cope’s rival Othniel Charles Marsh named another new species, Claosaurus annectens. During this period most hadrosaur fossils were being named as other earlier established generas such as Trachodon, Diclonius, Claosaurus and Hadrosaurus. Not all palaeontologists were in agreement as to what was and was not valid however, and published opinions varied greatly between them.
The first establishment of the name Edmontosaurus came in 1917 when Canadian palaeontologist Lawrence Lambe who at the time was naming two partially preserved individuals recovered from the Edmonton Formation (today known as the Horseshoe Canyon Formation). Lambe established the type species as Edmontosaurus regalis, and noted how the two specimens appeared to be similar to another species named Diclonius mirabilis.
In the 1920s further hadrosaur species such as Thespesius saskatchewanensis and Thespesius edmontoni were named. However, by the 1930s palaeontologists were starting to take note that many of the older species assigned to early genera did not quite fit in well where they were. Ultimately in 1942, Richard Lull and Nelda Wright created an entirely new genus called Anatosaurus which was based upon Claosaurus annectens, and also included other species such as Thespesius edmontoni, T. saskatchewanensis and Trachodon longiceps. This led to the Anatosaurus genus being represented by the species A. annectens (type), A. copei, A. edmontoni, A. longiceps, and A. saskatchewanensis.
Although essentially a wastebasket taxon for holding various species that just didn’t fit, Anatosaurus quickly became well known in popular science concerning dinosaurs. The name Anatosaurus literally translates as ‘duck lizard’ in reference to the duck-like bill that Anatosaurus would have had in life, and due to its popularity, the hadrosaurs subsequently became more commonly known as the ‘duck billed’ dinosaurs. During this time, almost every depiction of a duck-billed dinosaur was of an individual of Anatosaurus.
Towards the end of the twentieth century our understanding of hadrosaurs and particularly both Anatosaurus and Edmontosaurus was again changing. The trigger event of this was the studies of Michael K. Brett-Surman who was examining the Anatosaurus fossils as part of his graduate studies. Brett-Surman came to the conclusion that all of the Anatosaurus species with the exception of A. copei and A. longiceps, actually belonged to the older Edmontosaurus genus, though as a distinct species to the type species E. regalis. Although Brett-Surman’s papers were not capable of being officially recognised by the ICZN, they did get the attention of other palaeontologists who agreed with his reasoning. The type species of Anatosaurus, A. annectens (originally Claosaurus annectens), was used to create Edmontosaurus annectens, with all others except A. copei and A. longiceps additionally being assigned to the species.
Towards the end of the twentieth century our understanding of hadrosaurs and particularly both Anatosaurus and Edmontosaurus was again changing. The trigger event of this was the studies of Michael K. Brett-Surman who was examining the Anatosaurus fossils as part of his graduate studies. Brett-Surman came to the conclusion that all of the Anatosaurus species with the exception of A. copei and A. longiceps, actually belonged to the older Edmontosaurus genus, though as a distinct species to the type species E. regalis. Although Brett-Surman’s papers were not capable of being officially recognised by the ICZN, they did get the attention of other palaeontologists who agreed with his reasoning. The type species of Anatosaurus, A. annectens (originally Claosaurus annectens), was used to create Edmontosaurus annectens, with all others except A. copei and A. longiceps additionally being assigned to the species.
At the time of writing there are two established species of Edmontosaurus, the type species E. regalis and the later named E.annectens. Analysis of known fossils and their locations shows that both of these species lived in the same locations as one another but at different times, with E. regalis being recognised as a Campanian only era species, and E. annectens being a later Maastrichtian era species. This indicates that E.annectens replaced E. regalis as the dominant Edmontosaurus species.Edmontosaurus as a living dinosaur
Hadrosaurid dinosaurs can be divided into two main groups, lambeosaurines (with Lambeosurus as the type genus) which had hollow bony crests on their skulls, and saurolophines (with Saurolophus as the type genus) that had solid to no bony crests on their skulls. Edmontosaurus belongs to the latter group as the bone skull itself has no ornamental protuberances, though soft tissue display structures could have still been present, as indeed now seems the case.
As previously mentioned above, Edmontosaurus is represented by some of the most numerous and best preserved fossil remains known, which probably makes Edmontosaurus one of the best understood hadrosaurs. With that said however we must not let ourselves get over confident as it was only in 2013, almost a century after the genus was first named, that Edmontosaurus was discovered to have had a small soft tissue crest on the top of the skull. This crest was fairly small, situated on top of the skull above the eyes to the back of the skull, and has only been preserved as an impression on the rock surrounding the bones. It’s possible though that this feature might had been found sooner considering that in the very early days of palaeontology, collectors would only concentrate upon extracting fossil bones without looking for surrounding features. Indeed, many potential soft tissue impressions of such areas as skin and muscle tissue have been known to have been destroyed by collectors who only cared about bones.
The skull of Edmontosaurus in general was long and triangular, with the largest known complete skull being up to one hundred and eighteen centimetres long. Like other hadrosaurs, the front of the mouth would have had an additional covering of a keratinous beak. Usually keratin does not get preserved, but at least one specimen of Edmontosaurus has been so well preserved that we can measure that the keratin of the beak would have extended for at least eight centimetres in front of the mouth.
The overall form of the skull would change with age, with adults that had proportionately flatter and longer skulls than juveniles. This change in age is partially what contributed to some Edmontosaurus specimens being named as other species and genera, but when palaeontologists realised the age changes, the names were re-classified as synonyms to Edmontosaurus.
Many Edmontosaurus skulls are so well preserved that it has been possible for some researchers to make casts of the brain cavity. Relative to body size, the brain of Edmontosaurus was small in proportion, and elongate in form. Edmontosaurus are not expected to have been exceptionally intelligent since most of the brain was orientated towards primal functions such as sight and smell, but they were probably intelligent enough to survive on the landscape of late Cretaceous North America.
The teeth in the skull were arranged in the form of dental batteries in the posterior region of the mouth. The method of feeding for Edmontosaurus (and other hadrosaurs) is thought to have been along the lines of plants being cropped by the keratinous beak, and then ground by the teeth at the back. As in all dinosaurs, when these teeth became worn and lost, new teeth would erupt to replace them. A 2009 study by Williams et al studied the micro wear patterns on the teeth and concluded that Edmontosaurus were probably grazers as opposed to selective browsers.
As far as the rest of the body is concerned, Edmontosaurus were quite generic in their appearance. The centre of balance in the body would have been just in front of the hips, with the large rear legs serving the focus of the weight bearing function. Because of the developed rear legs, it certainly would have been possible for Edmontosaurus to adopt a bipedal posture, however they mostly seem to have been bipedal. The main evidence for this comes from the simple observation of the fore limbs which although are not as well developed as the rear limbs, are still formed for a weight bearing function. Fossil track ways of hadrosaurs also show that these kinds of dinosaurs walked about on all fours, and not just their rear legs.
Much analysis has been done upon how Edmontosaurus moved, with the 2009 study by Sellers et al being one of the better known. Hadrosaurs like Edmontosaurus were long thought to by earlier palaeontologists to have tried to escape from predators by running on their hind legs, but computer modelling from that study revealed that Edmontosaurus would have likely been faster if moving on all fours in the form of a gallop rather than running on just the rear legs.
As in other hadrosaurs, the tail vertebrae in the tail of Edmontosaurus were reinforced by a lattice network of tendons that became ossified with age. This means that the tail would have been quite rigid and not open to much movement save for at the base. Such a tail is speculated to have primarily been as a counterbalance, particularly when changing between quadrupedal and bipedal postures. Another function of the tail may have been as a display feature, perhaps being differently coloured than the rest of the body, though we cannot know this for certain at this time.
Preserved skin impressions have revealed that Edmontosaurus had non overlapping scales, most of which were small at being between one and three millimetres across. Larger polygonal scales ranging from below five to up to ten millimetres across are known to have been on areas such as the fore arms and shoulder. The smaller scales were more typical of the flanks while scales got slightly larger further up and down the body. There also seems to have been a frilly ridge of soft tissue that ran down the centreline of the neck and back. Preserved segments of this frill were about five centimetres long and about eight centimetres high.
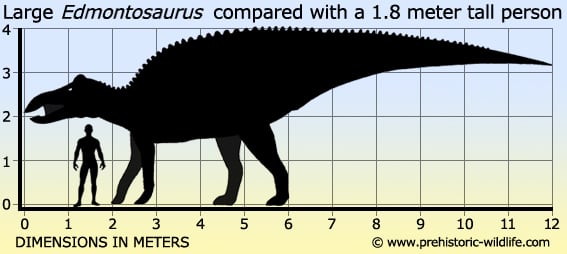
It used to be that how big an Edmontosaurus was depending upon which species you were talking about. The type species E. regalis has been considered to have been the larger with individuals reaching nine meters at adulthood with the oldest individuals reaching twelve and potentially even thirteen meters in length. The second species E. annectens has often been credited as being much smaller at about eight to nine meters long, however new studies of existing Edmontosaurus material has now revealed that E. annectens may have actually reached sizes up to twelve meters long as well. This would mean that E. anenctens was actually comparable to E. regalis in terms of size after all.
In terms of behaviour Edmontosaurus have been perceived to be herding animals based largely upon the evidence of bone beds of Edmontosaurus individuals collected together, ranging from a few, to many thousands of individuals. Herding would make sense for a plant eating animal to survive, especially an animal that lived on a landscape which included predators as large as itself, particularly tyrannosaurs such as Albertosaurus, Daspletosaurus and of course Tyrannosaurus itself.
There is also strong evidence to suggest that Edmontosaurus were frequently targeted and eaten by the predators of the North American late Cretaceous. Tyrannosaur tooth marks from Albertosaurus and Daspletosaurus are known upon Edmontosaurus fossils, though these can be perceived as signs of scavenging. Teeth marks from smaller theropod dinosaurs are also known upon the jaws of Edmontosaurus, suggesting that the neck may have been the preferred vulnerable spot targeted by such predators.
The most exciting discovery showing predator prey interaction though can be seen upon the damaged caudal vertebrae of a mounted Edmontosaurus specimen that is on display at the Denver Museum of Nature and Science. Some of the vertebrae on this specimen have been bitten into by a large predatory theropod with a bite pattern similar to what you would expect from a tyrannosaur. In addition to this the only large theropod that is known to have existed in the same formation as this individual Edmontosaurus and also to have been big enough to reach high enough to inflict such a bite is Tyrannosaurus.
What is really astonishing about this individual Edmontosaurus is that after the bite was inflicted, the bone started to heal, indicating that the attack was not immediately successful in killing it, and that the individual survived for long enough for the bone to partially heal. This proves that this Edmontosaurus was alive at the time of the attack and therefore it cannot be a case of scavenging. Further study of the specimen by renowned palaeontologist Kenneth Carpenter has also revealed that the left hip also has a partially healed fracture from an injury that likely happened before the attack that resulted in the tail injury. This may mean that the when this individual Edmontosaurus was attacked, it may have had a slight limp which resulted in it being targeted by a tyrannosaur.
Edmontosaurus is known to have had one of the broadest geographical distributions of all the known hadrosaurs, something that a 2008 study by Bell and Snively considered to have been the result of migratory behaviour. Migratory behaviour may have also been possible considering that if Edmontosaurus did live in groups, they may have had to continually move to avoid exhausting available food resources. In addition the occurrence of vast bone beds may have been caused by many Edmontosaurus drowning together as they tried to cross rivers swollen with flood water, as has been proposed for other types of dinosaur such as ceratopsians.
However there are doubts as to whether Edmontosaurus were truly migratory, potentially moving many hundreds if not thousands of miles with the seasonal growth of plants, or if they instead did not roam as far. One study by Chisamy et al in 2012, revealed that hadrosaur remains recovered from what were more polar regions, were from populations that lived there all the time rather than from a migratory population. Perhaps a more general scenario would be that as with most animals, Edmontosaurus stayed where the food was, only moving on to the next horizon for food when they had to.
Further Reading
– Supplement to the synopsis of the extinct Batrachia and Reptilia of North America – American Philosophical Society, Proceedings 12 (86): 41–52. – Edward Drinker Cope – 1871. – Report on the vertebrate paleontology of Colorado – U.S. Geological and Geographical Survey of the Territories Annual Report 2: 429–454. – Edward Drinker Cope – 1874. – Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado – U.S. Geological and Geographical Survey of the Territories Annual Report 1: 9–28. – Edward Drinker Cope – 1874. – On the characters of the skull in the Hadrosauridae – Proceedings of the Philadelphia Academy of Natural Sciences 35: 97–107. – Edward Drinker Cope – 1883. – The genus and species of the Trachodontidae (Hadrosauridae, Claosauridae) Marsh – Annals of the Carnegie Museum 1 (14): 377–386. John B. Hatcher – 1902. – The dinosaur Trachodon annectens – Smithsonian Miscellaneous Collections 45: 317–320 – Frederic A. Lucas – 1904. – The epidermis of an iguanodont dinosaur – Science 29 (750): 793–795. – Henry Fairfield Osborn – 1909. – Integument of the iguanodont dinosaur Trachodon – Memoirs of the American Museum of Natural History 1: 33–54 – Henry Fairfield Osborn – 1912. – The manus in a specimen of Trachodon from the Edmonton Formation of Alberta – The Ottawa Naturalist 27: 21–25. – Lawrence M. Lambe – 1913. – On the fore-limb of a carnivorous dinosaur from the Belly River Formation of Alberta, and a new genus of Ceratopsia from the same horizon, with remarks on the integument of some Cretaceous herbivorous dinosaurs – The Ottawa Naturalist 27: 129–135. – Lawrence M. Lambe – 1914. – On the genus Trachodon – Science 41 (1061): 658–660 – Charles W. Gilmore – 1915. – A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta. – The Ottawa Naturalist 31 (7): 65–73. – Lawrence M. Lambe – 1917. – The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta. Memoir 120. – Department of Mines, Geological Survey of Canada. pp. 1–79. – Lawrence M. Lambe – 1920. – A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta. Bulletin 38. – Department of Mines, Geological Survey of Canada. pp. 13–26. – Charles W. Gilmore – 1924. – Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper 40. – Geological Society of America. – Richard Swann Lull & Nelda E. Wright – 1942. – A reconsideration of the paleoecology of the hadrosaurian dinosaurs – American Journal of Science 262 (8): 975–997. – John H. Ostrom – 1964. – The posture of hadrosaurian dinosaurs – Journal of Paleontology 44 (3): 464–473. – Peter M. Galton – 1970. – Hadrosaurian dinosaur bills — morphology and function – Contributions in Science (Los Angeles County Museum of Natural History) 193: 1–14. – William J. Morris – 1970. – The evolution of cranial display structures in hadrosaurian dinosaurs – Paleobiology 1 (1): 21–43. – James A. Hopson – 1975. – A “segmented” epidermal frill in a species of hadrosaurian dinosaur – Journal of Paleontology 58 (1): 270–271. – John R. Horner – 1984. – Evidence of predatory behavior by theropod dinosaurs – Gaia 15: 135–144. – Kenneth Carpenter – 1998/2000. – Taphonomic aspects of theropod tooth-marked bones from an Edmontosaurus bone bed (Lower Maastrichtian), Alberta, Canada – Journal of Vertebrate Paleontology 19 – Aase Roland Jacobson, Michael J. Ryan – 2000. – Dinosaur forebrains – Journal of Vertebrate Paleontology 21 (3, Suppl.): 64A – Harry J. Jerison, John R.Horner& Celeste C. Horner – 2001. – New skin structures from a juvenile Edmontosaurus from the Late Cretaceous of North Dakota – Abstracts with Programs — Geological Society of America 35 (2): 13. – Tyler R. Lyson, Douglas H. Hanks & Emily S. Tremain – 2003. – An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD – Journal of Vertebrate Paleontology 23 (3, suppl.): 56A–57A. – Rebecca Gould, Robb Larson & Ron Nellermoe – 2003. – Microscale δ18O and δ13C isotopic analysis of an ontogenetic series of the hadrosaurid dinosaur Edmontosaurus: implications for physiology and ecology – Palaeogeography, Palaeoclimatology, and Palaeoecology 206 (2004): 257–287 – Kathryn J. Stanton Thomas & Sandra J. Carlson – 2004. – Preliminary depositional model for an Upper Cretaceous Edmontosaurus bonebed – Journal of Vertebrate Paleontology 26 (3, suppl.): 49A., Arthur Chadwick, Lee Spencer & Larry Turner 2006. – How to make a fossil: part 2 – Dinosaur mummies and other soft tissue. – The Journal of Paleontological Sciences. – Kenneth Carpenter – 2007. – A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses – Palaeontologia Electronica 11 (2) – Natalia Rybczynski, Alex Tirabasso, Paul Bloskie, Robin Cuthbertson, Casey Holliday – 2008. – Polar dinosaurs on parade: a review of dinosaur migration – Alcheringa 32 (3): 271–284. – Phil R. Bell & E. Snively – 2008. – Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids – Journal of Vertebrate Paleontology 28 (4): 1073–1088. – Casey M. Holliday & Lawrence M. Witmer – 2008. – Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA) – Proceedings of the Royal Society B 276 (1672): 3429–3437 – Phillip L. Manning, Peter M. Morris, Adam McMahon, Emrys Jones, Andy Gize, Joe H. S.Macquaker, G. Wolff, Anu Thompson, Jim Marshall, Kevin G. Taylor, Tyler Lyson, Simon Gaskell, Onrapak Reamtong, William I. Sellers, Bart E. van Dongen, Mike Buckley, Roy A. Wogelius – 2009. – Cranial variation in Edmontosaurus (Hadrosauridae) from the Late Cretaceous of North America – North American Paleontological Convention (NAPC 2009): Abstracts, p. 95a. – N. E. Campione – 2009. – Virtual palaeontology: gait reconstruction of extinct vertebrates using high performance computing -Palaeontologia Electronica 12 (3). – W. I. Sellers, P. L. Manning, T. Lyson, K. Stevens & L. Margetts – 2009. – Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding – Proceedings of the National Academy of Sciences 106 (27): 11194–11199. – Vincent S. Williams, Paul M. Barrett & Mark A. Purnell – 2009. – Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae): Implications for Latest Cretaceous Megaherbivore Diversity in North America. – PLoS ONE, 6(9): e25186 – N. E. Campione & D. C. Evans – 2011. – Complex dental structure and wear biomechanics in hadrosaurid dinosaurs – Science 338 (6103): 98–101 – Gregory M. Erickson, Brandon A. Krick, Matthew Hamilton, Gerald R. Bourne, Mark A. Norell, Erica Lilleodden & W. Gregory Sawyer – 2012. – Kinetic limitations of intracranial joints in Brachylophosaurus canadensis and Edmontosaurus regalis (Dinosauria: Hadrosauridae), and their implications for the chewing mechanics of hadrosaurids – The Anatomical Record 295 (6): 968–979. – Robin S. Cuthbertson, Alex Tirabasso, Natalia Rybczynski & Robert B. Holmes – 2012. – Hadrosaurs were perennial polar residents – The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. – A. Chinsamy, D. B. Thomas, A. R. Tumarkin-Deratzian & A. R. Fiorillo – 2012. – A Mummified Duck-Billed Dinosaur with a Soft-Tissue Cock’s Comb – Current Biology. – Phil R. Bell, Federico Fanti, Philip J. Currie & Victoria M. Arbour – 2013. – Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. – PLOS ONE. 12 (4). – H. Xing, J. C. Mallion & M. L. Currie – 2017. Skeletal Trauma with Implications for Intratail Mobility in Edmontosaurus annectens from a Monodominant Bonebed, Lance Formation (Maastrichtian), Wyoming USA. – PALAIOS. 35 (4): 201–214 – Bethania C. T. Siviero, Elizabeth Rega, William K. Hayes, Allen M. Cooper, Leonard R. Brand & Art V. Chadwick – 2020.









